

Содержание
Транспорт воды в растении
Транспорт воды в растении


Транспорт у растений
Клетки обмениваются различными веществами с окружающей их средой в результате диффузии. Однако перенос веществ обычной диффузией на большие расстояния неэффективен; возникает необходимость в специализированных системах транспорта. Такой перенос из одного места в другое осуществляется за счёт разности давлений в этих местах. Все переносимые вещества движутся с одинаковой скоростью в отличие от диффузии, где каждое вещество движется со своей скоростью в зависимости от градиента концентрации.
У животных можно выделить четыре основных типа транспорта: пищеварительную, дыхательную, кровеносную и лимфатическую системы. Часть из них были описаны ранее, к другим мы перейдем в следующих параграфах.
У сосудистых растений передвижение веществ осуществляется по двум системам: ксилеме (вода и минеральные соли) и флоэме (органические вещества). Передвижение веществ по ксилеме направлено от корней к надземным частям растения; по флоэме питательные вещества движутся от листьев.
Одним из важнейших механизмов транспорта веществ в растении является осмос. Осмос – это переход молекул растворителя (например, воды) из областей с более высокой концентрацией в области с более низкой концентрацией через полупроницаемую мембрану. Этот процесс похож на обычную диффузию, но протекает быстрее. Численно осмос характеризуется осмотическим давлением – давлением, которое нужно приложить, чтобы предотвратить осмотическое поступление воды в раствор.
В растениях роль таких полупроницаемых мембран играют плазматическая мембрана и тонопласт (мембрана, окружающая вакуоль). Если клетка контактирует с гипертоническим раствором (то есть раствором, в котором концентрация воды меньше, чем в самой клетке), то вода начинает выходить из клетки наружу. Этот процесс называется плазмолизом . Клетка при этом сморщивается. Плазмолиз обратим: если такую клетку поместить в гипотонический раствор (с более высоким содержанием воды), то вода начнёт поступать внутрь, и клетка снова набухнет. При этом внутренние части клетки (протопласт) оказывают давление на клеточную стенку. У растительной клетки набухание останавливается жесткой клеточной стенкой. У животных клеток жёстких стенок нет, а плазматические мембраны слишком нежны; необходим особый механизм, регулирующий осмос.
Еще раз подчеркнём, что осмотическое давление – величина скорее потенциальная, чем реальная. Она становится реальной только в отдельных случаях – например, при её измерении. Также необходимо помнить, что вода движется в направлении от более низкого осмотического давления к более высокому.
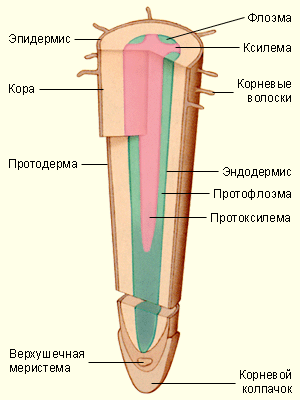

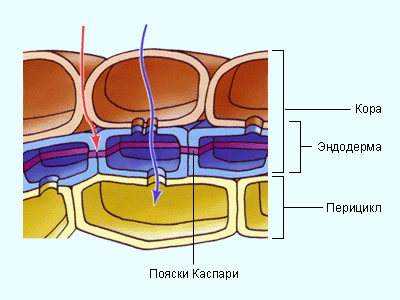
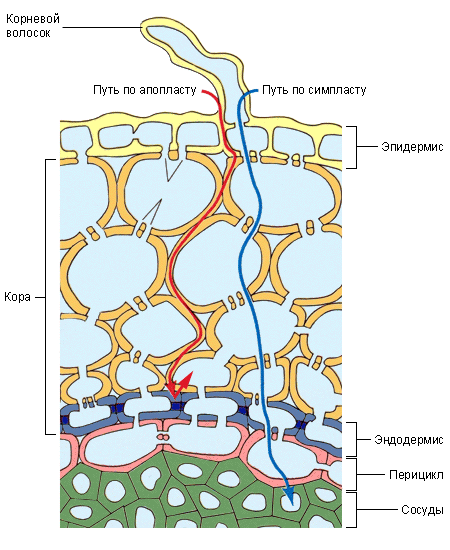
Основная масса воды поглощается молодыми зонами корней растений в области корневых волосков – трубчатых выростов эпидермиса. Благодаря им значительно увеличивается всасывающая воду поверхность. Вода поступает в корень за счёт осмоса и движется вверх к ксилеме по апопласту (по клеточным стенкам), симпласту (по цитоплазме и плазмодесмам), а также через вакуоли. Надо заметить, что в клеточных стенках имеются полоски, называемые поясками Каспари . Они состоят из водонепроницаемого суберина и препятствуют продвижению воды и растворённых в ней веществ. В этих местах вода вынуждена проходить через плазматические мембраны клеток; полагают, что таким образом растения защищаются от проникновения токсичных веществ, патогенных грибов и т. п.
Подъём воды по ксилеме происходит, по-видимому, за счёт испарения воды в листьях. В процеcсе испарения в кроне образуется недостаток воды. Поверхностное натяжение в сосудах ксилемы способно тянуть вверх весь столб воды, создавая массовый поток. Скорость подъёма воды составляет около 1 м/ч (до 8 м/ч в высоких деревьях); чтобы поднять воду к вершине высокого дерева, требуется давление порядка 40 атм. Следует иметь в виду, что одни только капиллярные эффекты способны поднять воду на высоту не более 3 м.
Вторая важная сила, участвующая в подъёме воды, – это корневое давление . Оно составляет 1–2 атм (в исключительных случаях – до 8 атм). Этой величины, конечно, недостаточно, чтобы в одиночку обеспечить движение жидкости, но её вклад у многих растений несомненен.
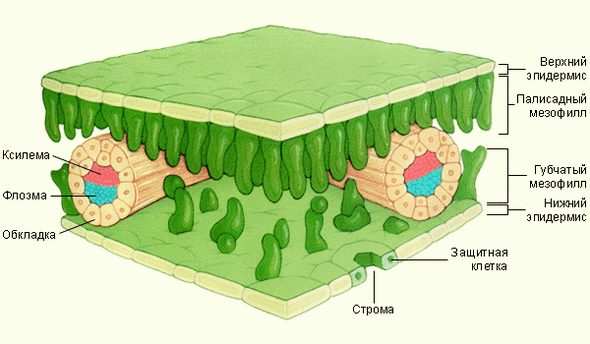
Попадая по ксилеме в листья, вода и минеральные вещества распределяются через разветвлённую сеть проводящих пучков по клеткам. Движение по клеткам листа осуществляется, как и в корне, тремя способами: по апопласту, симпласту и вакуолям. На свои нужды растение использует менее 1 % поглощаемой им воды, остальное в конце концов испаряется через восковый слой на поверхности листьев и стеблей – кутикулу (около 10 % воды) – и особые поры – устьица (90 % воды). Травянистые растения теряют в день около литра воды, а у больших деревьев эта цифра может доходить до сотен литров. Испарение воды ( транспирация ) осуществляется за счёт энергии солнца. Проще всего транспирацию наблюдать, если накрыть растение в горшке колпаком; на внутренней поверхности колпака будут собираться капельки жидкости.

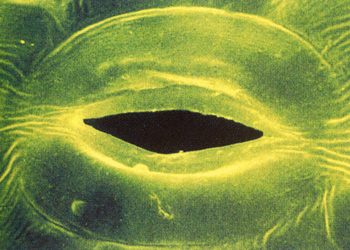
На скорость испарения влияют многие факторы; как внешние условия (свет, температура, влажность, наличие ветра, доступность воды в почве), так и особенности строения листьев (площадь поверхности листа, толщина кутикулы, количество устьиц). Ряд внешних факторов приводит к уменьшению диффузии воды из листьев, другие (например, отсутствие света или сильный ветер) вызывают замыкание устьиц (благодаря работе особых замыкающих клеток). Растения засушливых регионов имеют специальные приспособления для уменьшения транспирации: погруженные глубоко в листья устьица, густое опушение из волосков или чешуек, толстый восковой налёт, превращение листьев в колючки или иглы и другие. Осенний листопад в умеренных широтах также призван уменьшить испарение воды, когда наступят холода.

Из почвы растение получает не только воду, но и минеральные соли. Эти вещества движутся в корнях под действием диффузии. За счёт энергии дыхания возможен также их активный транспорт против градиента концентрации. Попадая в ксилему, минеральные вещества разносятся по всему организму с массовым током воды. Основным потребителем этих веществ являются растущие части растения.
Некоторые минеральные вещества, выполнив свою полезную функцию, могут перемещаться дальше вверх или вниз по флоэме. Это происходит, например, перед сбрасыванием листьев, когда накопленные листьями полезные вещества сохраняются, откладываясь в других частях растения.
У многоклеточных растений есть ещё одна транспортная система, предназначенная для распределения продуктов фотосинтеза, – флоэма. В отличие от ксилемы, органические вещества могут транспортироваться по флоэме и вверх, и вниз. 90 % переносимых веществ составляет сахароза, которая практически не участвует в метаболизме растения непосредственно и поэтому является идеальным углеводом для транспорта. Скорость движения сахара обычно составляет 20–100 см/ч; за день по стволу большого дерева может пройти несколько килограммов сахара (в сухой массе).
Каким образом столь большие потоки питательных веществ могут протекать в тонких ситовидных трубках флоэмы (их диаметр не превышает 30 мкм), не совсем понятно. По-видимому, вещества по флоэме распространяются массовым током, а не диффузией. Возможными механизмами транспорта являются обычное давление или электроосмос.

При повреждении флоэмы ситовидные трубки закупориваются в результате отложения каллозы на ситовидных пластинках. Безвозвратная утечка питательных веществ обычно прекращается уже через несколько минут после повреждения.
Транспорт воды в растении и его регуляция
Транспорт воды как важнейшая физиологическая функция. Вода является универсальной внутренней средой, в которой разыгрываются жизненные процессы. Она является, по образному выражению А. Сент-Дьёрдьи, «матрицей» жизни. Вместе с тем, содержащаяся в организме вода находится в постоянном движении, то есть живой организм подобен не изолированному водоему со стоячей водой, а проточному водоему, в котором вода непрерывно обменивается. Именно благодаря безостановочной циркуляции водных растворов реализуются взаимосвязи между отдельными клетками, тканями, органами (в том числе доставляются метаболиты и удаляются токсины); возникает круговорот веществ; поддерживается гомеостаз и функционирование организма как единого целого.
Пути транспорта воды в растении. Общепризнанно, что у животных циркуляция воды осуществляется по системам крово- и лимфообращения и эндогенно регулируется. Моторную роль при этом выполняют контрактильные системы, прежде всего сердце, остановка которого мгновенно приводит к смерти. Транспортные системы животных интенсивно изучаются и уже достигнут известный прогресс в понимании соответствующих движущих сил. К сожалению, в отношении растений этого сказать нельзя. Природа транспорта воды у них изучена несравненно слабее.
Главным транспортным путем воды у наземного растения считается восходящий ток. Обычно только его и имеют в виду, когда заходит речь о межклеточном (ближнем, по неспециализированным тканям) или дальнем (по проводящим тканям) транспорте воды. До сих пор распространено упрощенноепредставление, согласно которому восходящий водный ток своим происхождением обязан не столько потребности самого растения в передвижении по нему воды, сколько специфическим условиям существования растения. А именно: растение не имеет прочных покровов (как у животных), поэтому, находясь в наземно-воздушной среде жизни, согласно законов физики должно непрерывно испарять влагу в окружающую среду в процессе транспирации и одновременно восполнять свой водный запас за счет поглощения воды из почвы. То есть в сущности, растение выполняет всего-навсего роль канала между почвой и атмосферой. Но это не совсем так.
Не следует забывать, что в наземном растении, помимо восходящего, имеется ещё и нисходящий водный ток, отнюдь не зависящий от транспирационной тяги. Этот ток называется флоэмным. Формируется он в листьях за счет воды, поступающей с восходящим током, а затем возвращается в корни, доставляя в них ассимиляты и другие продукты жизнедеятельности листьев. Таким образом, в растении одновременно функционируют два противоположно направленных водных потока. Причем, становятся все более очевидными их структурные и функциональные взаимосвязи (Курсанов, 1976). Следовательно, во-первых, у растений существует круговорот водных растворов, а, во-вторых, имеется единая гидродинамическая система целого растения (Жолкевич и др., 1989) или единая транспортная система (Гамалей, 1997).
Доказательством этого является то, что циркуляция водных растворов обнаружена (в опытах с тритиевой водой) у вообще ничего не испаряющих, полностью погруженных в воду высших растений и водорослей. Наряду с этим установлено, что в наземном растении восходящий водный ток возможен при практически полностью подавленной транспирации, то есть в отсутствие транспирационной тяги. Все это ставит под сомнение правомерность представления о возникновении и поддержании восходящего водного тока лишь вследствие необходимости компенсировать неизбежные, но не особенно нужные растению потери влаги при транспирации, а о самом этом токе как о простом физическом явлении.
Ниже остановимся лишь на части единой гидродинамической системы целого растения – восходящем водном токе. Рассмотрим этапы восходящего водного тока. Начнем с радиального транспорта воды от поверхности корня до сосудов центрального цилиндра.
Механизм передвижения воды по растению
Вода в растении, преодолевая силы гравитации, должна подниматься на всю его высоту (эвкалипт, секвойя — до 140 м, береза — до 25 м. дуб — до 40 м, сосна и ель — до 50 м). К этому следует добавить и значительную протяженность водопроводящей системы корней.
Восходящий поток воды в растении идет по сосудам ксилемы, лишенным цитоплазмы. Помимо работы нижнего концевого двигателя и присасывающего действия транспирации (верхний концевой двигатель) в передвижении воды по капиллярным сосудам ксилемы участвуют силы когезии — сцепление молекул воды друг с другом и силы адгезии сцепление молекул воды со стенками сосудов. Обе силы препятствуют также образованию пузырьков воздуха, способных закупорить сосуд. В зависимости от анатомического строения древесины линейная скорость восходящего тока колеблется от 1 — 6 м/ч у хвойных и рассеянно-сосудистых древесных пород, до 25 — 60 м/ч у кольцесосудистых. Такая скорость зарегистрирована летом в полдень. Скорость передвижения воды по растению в течение суток изменяется и в основном соответствует интенсивности транспирации.
Вода из клеток листа и непосредственно из сосудов ксилемы поступает во флоэмные окончания по осмотическому градиенту, возникающему вследствие накопления в клетках флоэмы сахаров и других органических соединений, которые образуются в клетках листьев и переносятся в клетки флоэмы в результате активной работы транспортных насосов. Нисходящий флоэмный ток доставляет органические соединения тканям корня, где они используются в метаболизме. В корне окончания проводящих пучков флоэмы, как и в листе, располагаются вблизи элементов ксилемы и вода по осмотическому градиенту поступает в ксилему и движется вверх с восходящим током. Таким образом происходит обмен воды в проводящей системе корня и листьев.
Транспирация
1. Характеристика и значение транспирации.
Транспирация — это физиологический процесс испарения воды растением. К.А. Тимирязев назвал транспирацию «необходимым физиологическим злом». Из поступившей в растение воды 99,8 % теряется в результате транспирации и лишь 0,02 % используется на метаболические процессы, поддержание тургора и в транспорте органических соединений по флоэме.
Основным органом транспирации является лист. Из-за необходимости максимального контакта с воздушной средой растение имеет очень большую листовую поверхность. Увеличение поверхности облегчает поглощение СО2, улавливание света и это же создает огромную поверхность испарения. В результате потери воды клетками листьев в них снижается водный потенциал, т. е. возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из ксилемы жилок и передвижению воды по ксилеме из корней в листья.
Таким образом, верхний концевой двигатель, обеспечивающий передвижение воды вверх по растению, создается и поддерживается высокой сосущей силой транспирирующих клеток листовой паренхимы. Поэтому сила верхнего концевого двигателя будет тем больше, чем активнее транспирация. Верхний концевой двигатель может работать при полном отключении нижнего концевого двигателя, причем для его работы используется не метаболическая энергия, а энергия внешней среды — температура и движение воздуха.
Различают устьичную и кутикулярную транспирацию. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом и определяется толщиной слоя кутикулы. Однако у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды. Трещины в кутикуле могут появляться и после временного завядания листьев, благодаря чему транспирация усиливается. Кутикулярная транспирация зависит от оводненности листа. При насыщении кутикулы водой испарение идет интенсивнее, а при подсыхании кутикулы — снижается. В целом, кутикулярная транспирация меньше зависит от условий внешней среды по сравнению с устьичной.
Некоторое количество воды выделяется в результате транспирации почек, поскольку почечные чешуи полностью не предохраняют их от потери влаги. Репродуктивные органы также теряют воду и в некоторых случаях эти потери могут быть очень значительными: например, корзинки подсолнечника, коробочки мака и плоды перца транспирируют сильнее, чем листья данных растений в тех же условиях. Кроме того, вода испаряется с поверхности ветвей и стволов древесных растений через чечевички и окружающие их слои пробки — перидермалышя транспирация. Хотя общее количество воды, испаряемой через чечевички, значительно меньше того, которое теряется через листья, фактически интенсивность транспирации на единицу испаряющей поверхности нередко мало различается в обоих случаях. Вследствие транспирации ветвей в зимнее время часто возникает водный дефицит и растения гибнут в результате обезвоживания.
1. Транспирация повышает сосущую силу испаряющих клеток и создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
2. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества от корней к жизненно важным частям растения. Чем интенсивнее транспирация, тем быстрее идет этот процесс.
3. Защищает растение от перегрева прямыми солнечными лучами; температура свежего листа может быть на 7 °С ниже температуры завядающего слаботранспирирующего листа.
4. Препятствует полному насыщению клеток водой, способствуя тем самым оптимизации целого ряда процессов метаболизма, так как при небольшом водном дефиците (до 5 %) активность синтетаз и интенсивность фотосинтеза оказываются наибольшими.
Дата добавления: 2018-02-28 ; просмотров: 933 ; ЗАКАЗАТЬ РАБОТУ
Новое в блогах
Движение воды по растению
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух концевых двигателей водного тока: верхнего концевого двигателя, или присасывающей силы испарения (транспирации), и нижнего концевого двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является присасывающая сила транспирации, в результате которого возникает градиент водного потенциала. Водный потенциал – это мера энергии, используемой водой для передвижения. Водный потенциал и сосущая сила одинаковы по абсолютному значению, но противоположны по знаку. Чем меньше насыщенность водой данной системы, тем меньше (более отрицателен) ее водный потенциал. При потере воды растением в процессе транспирации создается ненасыщенность клеток листа водой, как следствие, возникает сосущая сила (водный потенциал падает). поступление воды идет в сторону большей сосущей силы, или меньшего водного потенциала.
Таким образом, верхний концевой двигатель водного тока в растении – это присасывающая сила транспирации листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Кроме верхнего концевого двигателя водного тока, в растениях существует нижний концевой двигатель. Это хорошо доказывается на примере таких явлениях, как гуттация.
Листья растений, клетки которых насыщены водой, в условиях высокой влажности воздуха, препятствующей испарению, выделяют капельно-жидкую воду с небольшим количеством растворенных веществ – гуттация. Выделение жидкости идет через специальные водные устьица – гидаторы. Выделяющаяся жидкость – гутта. Таким образом, процесс гуттации является результатом одностороннего тока воды, происходящего в отсутствие транспирации, и, следовательно, вызывается какой-то иной причиной.
К такому же выводу можно прийти и при рассмотрении явления плач растений. Если срезать побеги растения и к срезанному концу присоединить стеклянную трубку, то по ней будет подниматься жидкость. Анализ показывает, что это вода с растворенными веществами – пасока. В некоторых случаях, особенно в весенний период, плач наблюдается и при надрезе веток растений. Определения показали, что объем выделяющейся жидкости (пасоки) во много раз превышает объем корневой системы. Таким образом, плач – это не просто вытекание жидкости в результате пореза. Все сказанное приводит к выводу, что плач, как и гуттация, связана с наличием одностороннего тока воды через корневые системы, не зависящего от транспирации. Силу, вызывающую односторонний ток воды по сосудам с растворенными веществами, не зависящую от процесса транспирации, называют корневым давлением. Наличие корневого давления позволяет говорить о нижнем концевом двигателе водного тока. Корневое давление можно измерить, присоединив манометр к концу, оставшемуся после срезания надземных органов растения, или поместив корневую систему в серию растворов различной концентрации и подобрав такую, при которой плач прекращается. Оказалось, что корневое давление равняется примерно 0,1 – 0,15 МПа (Д.А.Сабинин). Определения, проведенные советскими исследователями Л.В.Можаевой, В.Н.Жолкевичем, показали, что концентрация наружного раствора, останавливающего плач, значительно выше концентрации пасоки. Это позволило высказать мнение, что плач может идти против градиента концентрации. Было показано также, что плач осуществляется только в тех условиях, в которых нормально протекают все процессы жизнедеятельности клеток. Не только умерщвление клеток корня, но и снижение интенсивности их жизнедеятельности, в первую очередь интенсивность дыхания, прекращает плач. В отсутствии кислорода, под влиянием дыхательных ядов, при понижении температуры плач приостанавливается. Все сказанное позволило Д.А.Сабинину дать следующее определение: плач растений – это прижизненный односторонний ток воды и питательных веществ, зависящий от аэробной переработки ассимелятов. Д.А.Сабинин предложил схему, объясняющую механизм одностороннего тока воды в корне. Согласно этой гипотезе, клетки корня поляризованы в определенном направлении. Это проявляется в том, что в разных отсеках одной и той же клетки процессы обмена веществ различны. В одной части клетки идут усиленные процессы распада, в частности, крахмала на сахара, вследствие чего концентрация клеточного сока возрастает. На противоположном конце клетки преобладают процессы синтеза, благодаря чему концентрация растворенных веществ в этой части клетки уменьшается. Надо учитывать, что все эти механизмы будут работать только при достаточном количестве воды в среде и не нарушенном обмене веществ.
Согласно другой гипотезе, зависимость плача растений от интенсивности дыхания является косвенной. Энергия дыхания используется для поступления ионов в клетки коры, откуда они десорбируются в сосуды ксилемы. В результате концентрация солей в сосудах ксилемы повышается, что и вызывает поступление воды.
Передвижение воды по растению
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы. Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, — апопласта и симпласта. Апопласт – это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы. Симпласт – это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему. Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая – по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту. Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии. Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 1).

Рис. 1. Путь воды в растении.
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды. Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Видео: Движение воды и органических веществ по стеблю.
Источники:
http://www.ebio.ru/org12.html
http://studopedia.ru/13_36755_transport-vodi-v-rastenii-i-ego-regulyatsiya.html
http://studopedia.net/2_46481_mehanizm-peredvizheniya-vodi-po-rasteniyu.html
http://maxpark.com/community/4057/content/3267272

