

Содержание
- 1 Отдел аскомикота или сумчатые грибы краткое содержание
- 1.1 Лекции по фитопатологии — файл 1.doc
- 1.2 Отдел АСКОМИКОТА (ASCOMYCOTA), или СУМЧАТЫЕ ГРИБЫ
- 1.3 КЛАСС АРХЕАСКОМИЦЕТЫ — ARCHAEASCOMYCETES
- 1.4 (syn. TAPHRINOMYCETES)
- 1.5 Отдел сумчатые грибы, или Аскомицеты (Ascomycota)
- 1.6 ОТДЕЛ СУМЧАТЫЕ ГРИБЫ ИЛИ АСКОМИЦЕТЫ (ASCOMYCOTA);
- 1.7 ОТДЕЛ АСКОМИКОТА (ASCOMYCOTA), ИЛИ СУМЧАТЫЕ ГРИБЫ
Отдел аскомикота или сумчатые грибы краткое содержание
Лекции по фитопатологии — файл 1.doc
Доступные файлы (1):
- Смотрите также:
- Общее языкознание и теория межкультурной культурной коммуникации[ лекция ]
- Электронные лекции по истории древнего мира[ лекция ]
- по природопользованию[ лекция ]
- по социальной работе[ документ ]
- по Буровзрывным работам (для получения ЕКВ)[ лекция ]
- по энергосбережению[ документ ]
- Автомобили (конструкция)[ документ ]
- Процессы открытых горных работ[ лекция ]
- Экономическая география[ лекция ]
- по международному маркетингу[ лекция ]
- по вычислительной математике[ лекция ]
- История российской рекламы[ документ ]
Отдел АСКОМИКОТА (ASCOMYCOTA), или СУМЧАТЫЕ ГРИБЫ
Аскомицеты — одна из самых больших групп грибов, включающая 30 000 видов. Для сумчатых грибов характерен многоклеточный (септированный) мицелий. В состав их клеточной стенки входят полисахариды хитин и β-глюкан. как у хитридиомицетов, у части видов, объединяемых в класс Гемиаскомицеты -маннан и β-глюкан.
^ Основной признак грибов отдела — формирование в результате полового процесса асков с аскоспорами.
В цикле развития многих аскомицетов большое значение имеет бесполое размножение. Оно осуществляется при помощи конидий, которые образуются на гаплоидном мицелии экзогенно на конидиеносцах разного строения.
Конидиальное спороношение служит для массового расселения аскомицетов. Обычно конидии образуются на живых растениях, а аскоспоры (за немногими исключениями) — после отмирания растения или его частей в конце вегетации либо после перезимовки.
По месту формирования сумок, особенностям их строения и результатам молекулярно-генетических исследований во многих системах грибов отдел Аскомикота подразделяют на следующие классы:
- Археаскомицеты — Archaeascomycetes (syn.Taphrinomycetes);
- Гемиаскомицеты, Голосумчатые — Hemiascomycetes — плодовые тела отсутствуют, сумки прототуникатные, образующиеся непосредственно на мицелии или при слиянии одиночных клеток;
- Настоящие сумчатые — Ascomycetes (sуn. Еuascomycetes) — сумки эутуникатные — унитуникатные, реже прототуникатные, образуются внутри или на поверхности плодовых тел;
- Локулоаскомицеты — Loculoascomycetes (sуn. Dothideomycetes) сумки эутуникатные — битуникатные, образуются в особых полостях (локулах), возникающих в сплетении мицелия — аскостроме, или псевдотеции.
Аскомикота включает около 75% всех описанных видов грибов. В настоящее время к сумчатым грибам относят также несовершенные (анаморфные, митотические) грибы — дейтеромицеты (в традиционном смысле), характеризующиеся сходным строением вегетативных структур и клеточной стенки при отсутствии в цикле развития сумок и наличии только конидиалыюй (бесполой) стадии — анаморфы. В эту же группу включают лихенообразующие симбиотрофные грибы, составляющие почти четверть видов Аскомикота. В ряде систем грибов (в классических учебниках) и до настоящего времени лишайники рассматриваются в качестве самостоятельного отдела — Lichenes, или Мусорhycophyta.
Представители отдела Аскомикота

1 — здоровые плоды;
2 — пораженные плоды;
3 — сумки и аскоспоры
слева — проявление болезни на листьях дуба;справа — схема развития мучнисторосяного гриба. Мицелий (I) и конидиальное спороношение (2,3) развиваются на поверхности ткани; внутрь субстрата.в эпидермальные клетки, заходят только гаустории (4)


Колос со склероциями,

Белая гниль подсолнечника:
1-поражение стебля; а- прорастание склероция

а) пораженный лист и плод яблони
б) псевдотеций с сумками,
в) конидиеносцы с конидиями
КЛАСС АРХЕАСКОМИЦЕТЫ — ARCHAEASCOMYCETES
(syn. TAPHRINOMYCETES)
Класс выделен на основании сравнения результатов секвенирования нуклеиновых кислот. Это наиболее древняя группа грибов, являющаяся исходной для остальных Аскомикота. Сумки у представителей этого класса эутуникатные. Плодовые тела отсутствуют. Группа разнородна по морфологии: некоторые виды одноклеточные, другие образуют как одиночные клетки, так и многоклеточные гифы.
Практический интерес для фитопатологии представляют три порядка: Сахаромицетовые. Тафриновые и Протомицетовые. Порядок Сахаромицетовые включает в основном дрожжевые грибы — сапротрофы. В порядок Тафриновые объединены грибы, паразитирующие в растениях, в порядок Протомицетовые — развивающиеся на дикорастущих растениях.
^ Порядок Сахаромицетовые — Saccharomycetales . В порядок Сахаромицетовые объединяют формы, отличающиеся строением вегетативного тела. Мицелий у них или нитчатый с перегородками, непочкующийся, или вначале нитчатый, а затем распадающийся и почкующийся.
Зигота развивается при слиянии двух отростков мицелия или двух отдельных свободноживущих клеток. Аски образуются как одиночные клетки.
Большинство сахаромицетов — сапротрофы, обитающие на поверхности плодов и вегетативных частей растений, в нектаре цветков, в почве. Отдельные виды паразитируют на растениях. Так, Spermophthora gossypii встречается на волокне в нераскрытых коробочках хлопчатника, Endomyces mali — один из возбудителей гнили яблок.
Большое практическое значение имеет семейство Сахаромицетовые — Saccharomycetaceae, в котором представлены дрожжи — возбудители спиртового брожения и продуценты кормового белка.
^ Порядок Тафриновые — Taphrinales . В порядок Тафриновые входит одно семейство тафриновые – Taphrinaceae и один род тафрина — Taphrina.. Он объединяет около 100 видов грибов паразитов высших растений, вызывающих гипертрофию и деформации пораженных органов.
Порядок тафриновые (Taphrinales) являются облигатными паразитами с узкой специализацией, вызывают на высших растениях образований галлов, «ведьминых метел», деформацию листьев, различные уродства. Плодовые тела отсутствуют. Бесполое размножение у грибов этого рода отсутствует, аскоспоры способны почковаться. Аски этих грибов образуются непосредственно на мицелии плотным слоем, расположенным под кутикулой пораженных органов растения. Мицелий грибов располагается по межклетникам и в клетках тканях растений-хозяев. У многих видов мицелий зимующий, сохраняющийся из года в год в трещинках коры, в стеблях и почках пораженных растений.
Заражение происходит гаплоидными аскоспорами. Разрастаясь по тканям, гифы проникают между эпидермисом и кутикулой. Здесь происходит расчленение на двухъядерные клетки, в которых между ядрами происходит кариогамия. В верхней части клеток возникает гифальный вырост, в который перемещается диплоидное ядро. Постепенно вырост разрывает кутикулу и превращается в сумку, оказывающуюся на поверхности зараженного органа.
Тафриновые поражают в основном плодовые косточковые. Заболевания проявляются в нарушении нормального развития зараженных органов и в возникновении в них различных уродств.
Установлено, что возникновение деформаций у зараженных растений связано со способностью тафриновых грибов выделять вещества, вызывающие интенсификацию синтеза фитогормонов самими растениями-хозяевами. Тафриновые выделяют такие фитогормоны, как β-индолилуксусная кислота и вещества типа цитокининов, которые усиливают их синтез.
Среди возбудителей болезней персика наиболее опасным является тафрина деформирующая (T.deformans), распространенная почти по всему земному шару. Болезнь носит название курчавость листьев персика. Листья, зараженные грибом при распускании почек, недоразвиваются, увеличиваются почти в 2 раза по сравнению с незараженными, жилки остаются укороченными. Особенно сильно разрастаются те листья, которые находятся в нижней части однолетних побегов. Важно отметить, что возбудитель курчавости способен заразить только молодые листья.
Помимо листвы, деформирующая тафрина заражает стебли и плоды. Стебли при этом желтеют, искривляются и утолщаются. Плоды на зараженных побегах недоразвиваются и деформируются. У них появляются тканевые разрастания и они, как и листья, опадают.
Сливовая тафрина (T.pruni) поражает сливу, вишню, алычу, а также терн и черемуху. Ее гифы, возникающие при прорастании отпочковавшихся аскоспор, проникают через цветоножки в завязь и при благоприятных условиях — повышенной влажности и температуре 17-I9°C — во время цветения через две недели или через месяц после заражения вызывают появление уродливых плодов. У зараженных плодов (покрытых беловатым восковидным налетом — сумчатым спороношением) разрастается перикарпий, косточка и зародыш не образуются и вместо них возникает обширная полость.
^ Порядок Протомицетовые – Рrotomycetales . Среди грибов этого порядка много возбудителей болезней дикорастущих растений. Эндогенно формирующийся мицелий расположен в межклетниках тканей растения-хозяина. В мицелии образуются крупные шаровидные или продолговатые хламидоспоры с гладкой оболочкой. Хламидоспоры зимуют. Весной из них развиваются аски с аскоспорами. Аскоспоры после выхода из асков копулируют, затем прорастают ростком диплоидного мицелия, проникающим в ткань растения. Паразитические грибы порядка Протомицетовые стимулируют образование опухолей на стеблях, черешках листьев, жилках растений-хозяев.
В состав порядка входят следующие роды: протомицес — Protomyces, протомикопсис — Protomycopsis и волкартия — Volkartia. Наиболее распространенный паразит — Protomyces macrosporus, вызывающий на листовых черешках сныти образование желваков.
Отдел сумчатые грибы, или Аскомицеты (Ascomycota)
Этот обширный отдел грибов насчитывает около 30 000 видов. Строение и образ жизни их очень разнообразны. Характернейшим признаком сумчатых грибов является образование сумки (аск), имеющей всегда строго определенное число спор (8 или 4, реже 2). Редко сумки возникают непосредственно на мицелии; почти всегда они располагаются целым слоем, получившим название гимениального слоя или гимения, чередуясь с тонкими бесплодными нитями — парафизами (рис. 270, 2—3). Гимений находится на поверхности плодового тела или внутри него.
Плодовые тела у сумчатых грибов имеют различную форму и составлены плотным сплетением гиф вегетативного мицелия. Открытые плодовые тела, имеющие форму блюдец, получили название апотециев (рис. 270, 1—2). Полуоткрытые плодовые тела, имеющие вид кувшинчиков с узким горлышком, называются перитециями (рис. 270, 3); совершенно замкнутые плодовые тела называются клейстокарпиями (рис. 270, 4).

Образованию плодового тела нередко предшествует появление половых органов. У некоторых сумчатых сумка образуется на мицелии в результате слияния двух совершенно одинаковых клеток; зигота непосредственно превращается в сумку (дрожжи). В других случаях в начале развития сумчатых плодовых тел закладывается несколько пар половых органов, состоящих каждая из женского органа — архикарпа и мужского — антеридия (рис. 271, 1—3). Архикарп образован здесь двумя клетками: верхней цилиндрической, называемой трихогиной, и нижней шаровидной, называемой аскогоном. Антеридий состоит из одной цилиндрической клетки. Как антеридий, так и аскогон многоядерны, но содержимое их совершенно лишено какой-либо дифференцировки на отдельные гаметы. Оплодотворение происходит таким образом, что антеридий прикладывается к кончику трихогины и переливает в нее свое содержимое (содержимое трихогины к этому времени дегенерирует). Отсюда содержимое антеридия вместе с многочисленными ядрами переливается через отверстие, образующееся в перегородке у основания трихогины, в аскогон, и здесь формируются дикарионы (сближенные, но не слившиеся еще ядра), которые располагаются в постенном слое цитоплазмы. Сейчас же после этого из аскогона начинают развиваться мешковидные ветвящиеся выросты — аскогенные гифы (сумкородные гифы), куда переходят дикарионы. Оба ядра дикариона делятся одновременно и параллельно друг другу (рис. 271, 4). Затем аскогенные гифы разделяются перегородками на клетки, каждая из которых, в том числе и верхние, содержит всегда по одному дикариону. Сумки развиваются из аскогенных гиф таким образом: кончик верхней клетки аскогенной гифы загибается крючком, и оба ядра делятся одновременно. Из получившихся четырех ядер два (не сестринские) остаются в месте перегиба крючка; из других двух одно отходит в основание крючка, а другое в загнутый кончик. Затем появляются две перегородки, вычленяющие среднюю двуядерную клетку, которая и является материнской клеткой сумки (рис. 271, 4). Ее два ядра затем сливаются, и зигота делится трижды (первое деление редукционное) (рис. 272). В результате получается восемь аскоспор, свободно плавающих в сумке.

Аск, или сумка, является тем органом, где происходит половой процесс.
Цикл развития сумчатого гриба (сморчка) показан на рисунке 283.
Одновременно с описанным процессом развития аскогенных гиф сумки обрастают гифами мицелия, слагающими главную массу плодового тела. Аскогенные гифы поэтому не растут свободно, а заключены в это сплетение, и только образуемые ими сумки высовываются на его поверхность, располагаясь здесь слоем гимения.
Отдел сумчатых грибов разделяется на три подотдела, которые, в свою очередь, делятся на классы:
- Подотдел Pezizomycotina (Ascomycotina).
- Класс Артониомицеты (Arthoniomycetes),
ОТДЕЛ СУМЧАТЫЕ ГРИБЫ ИЛИ АСКОМИЦЕТЫ (ASCOMYCOTA);
Basidiomycota
Ascomycota
Zygomycota
Классы Ascomycetes, Neolectomycetes, Pneumocystidomycetes, Saccharomycetes, Schizosaccharomycetes, Taphrinomycetes/
Классы Basidiomycetes, Urediniomycetes, Ustilaginomycetes.
Сумчатые грибы, или аскомицеты,— один из крупнейших отделов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот отдел грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10—20 см (сморчки, строчки). Но все эти многообразные формы связаны общим происхождением и имеют ряд общих черт, на основании которых они объединяются в этот класс.
Основной признак аскомицетов — образование в результате полового процесса сумок (или асков) — одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8. Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах.
Вегетативное тело аскомицетов — разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. У некоторых аскомицетов мицелий может распадаться на отдельные клетки или почковаться. У дрожжей настоящего мицелия нет, а вегетативное тело представлено одиночными почкующимися, реже делящимися клетками, иногда образующими псевдомицелий.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды).
Конидиальные спороношения развиваются в период вегетации грибов и служат для их массового расселения. У аскомицетов — паразитов растений они обычно образуются на живом растении, а сумчатые спороношения, за немногими исключениями,— после отмирания растения или его частей, в конце периода вегетации или после перезимовки.
Половой процесс, типичный для аскомицетов,— гаметангиогамия, т. е. слияние двух гаметангиев — специализированных клеток, не дифференцированных на гаметы.
Для многих аскомицетов характерна морфологическая редукция полового процесса. У некоторых представителей этого класса антеридии отсутствуют или не функционируют. В этом случае их функции могут выполнять конидии, вегетативные гифы, а часто мелкие специализированные клетки, называемые спермациями.
В сумке аскомицетов обычно образуется 8 аскоспор. Однако наблюдаются многочисленные отклонения от типа. Форма аскоспор очень разнообразна — от шаровидных или эллипсоидальных до нитевидных. Аскоспоры могут быть одноклеточными или иметь поперечные перегородки, реже — поперечные и продольные перегородки (муральные споры). У некоторых аскомицетов аскоспоры имеют разнообразные придатки, играющие роль в их распространении.
По строению оболочки и функциям сумки аскомицетов делят на две большие группы — прототуникатные и эутуникатные. Прототуникатные сумки имеют тонкую недифференцированную оболочку, которая разрушается или растворяется, освобождая аскоспоры пассивно. Такая сумка служит только местом формирования спор, но не участвует активно в их распространении. Эутуникатные сумки характеризуются более плотными оболочками, часто со специальными приспособлениями для вскрывания сумки. Они активно участвуют в распространении аскоспор (активное выбрасывание спор из сумки). По строению оболочки эутуникатные сумки могут быть двух типов: унитуникатные и битуникатные. У унитуникатных сумок оболочка относительно тонкая и выглядит однослойной, на вершине сумок имеется обычно апикальный аппарат различного строения, служащий для их вскрывания. Битуникатные сумки имеют ясновыраженную двухслойную оболочку, состоящую из жесткого наружного и эластичного внутреннего слоев. При созревании аскоспор наружный слой оболочки разрушается, начиная с вершины; внутренний слой под действием повышенного тургорного давления растягивается, и происходит активное выбрасывание аскоспор.
Форма сумок округлая или овальная (у гемиаскомицетов и аскомицетов с замкнутыми плодовыми телами) или цилиндрическая (у аскомицетов с активным выбрасыванием аскоспор).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших — в специальных вместилищах — плодовых телах. Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции — настоящие плодовые тела, развитие которых происходит по аскогимениальному типу — образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.
Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах — плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела.
Аскомицеты широко распространены в природе во всех географических областях, на всех доступных для грибов субстратах. Они обитают как сапрофиты в почве, в лесной подстилке, на разнообразных растительных субстратах (древесина, отмершие растения и т. п.). Некоторые группы аскомицетов заняли своеобразные экологические ниши, недоступные для других грибов. Например, кератинофильные грибы, развивающиеся на различных субстратах животного происхождения, содержащих кератин. Некоторые аскомицеты обитают в морях или пресных водоемах на погруженной в воду древесине. Сапрофитные аскомицеты активно участвуют в минерализации органических веществ в природе, особенно в разложении растительных остатков, содержащих целлюлозу. Многие сапрофитные аскомицеты вызывают плесневение и порчу разнообразных материалов и изделий, а также пищевых продуктов.
Многочисленные аскомицеты паразитируют на различных организмах — на растениях (грибах, водорослях, лишайниках и высших), а также на животных и человеке, нередко вызывают серьезные заболевания.
Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
Типичные представители аскомицетов:
2. Тафрина (паразиты высших растений, вызывающие гипертрофию и деформацию пораженных органов).
3. Мучнисторосяные грибы (облигатные паразиты высших растений, вызывающие болезнь – мучнистая роса).
4. Спорыньевые (спорынья) — паразиты растений (злаков, осок).
5. Пецицевые (Пецица, Сморчки и Строчки). Встречаются на влажных почвах в лесах (на гниющей древесине).
КЛАСС БАЗИДИОМИЦЕТЫ (BASIDIOMYCETES)
Базидиомицеты — высшие грибы с многоклеточным мицелием. К ним относятся около 30 тыс. видов (и микроскопические грибы, и грибы с крупными плодовыми телами). Среди этих грибов есть паразиты растений (например, широко распространенные и очень опасные для сельскохозяйственных растений головневые и ржавчинные грибы), многочисленные почвенные сапрофиты — хорошо всем известные шляпочные грибы (например, шампиньоны, навозники). К базидиомицетам относятся и микоризообразующие шляпочные грибы, которые успешно развиваются только в тесном контакте с корнями древесных растений (например, белый, подберезовик, подосиновик и многие другие лесные грибы).
Есть среди базидиальных грибов и сапрофиты на древесине — это многочисленные трутовики — активные разрушители древесины и валежника.
Половое спороношение у них — базидиоспоры, т. е. экзогенные споры на особых выростах — базидиях. Такая базидия закладывается из двухъядерных клеток. Половых органов нет. Половой процесс осуществляется путем слияния двух вегетативных клеток гаплоидного мицелия, вырастающего из базидиоспоры и состоящего из одноядерных клеток. Базидии с базидиоспорами могут возникать прямо на мицелии. Но у большинства базидиомицетов они образуются на плодовых телах или внутри них. Рассеивание базидиоспор происходит путем их активного отбрасывания. В основе этого лежит повышение в самой базидии внутриклеточного давления в результате гидролиза гликогена. В дальнейшем она подхватывается токами воздуха. Плодовые тела базидиомицетов различны по форме и консистенции. Они могут быть паутинистыми, рыхлыми, плотно-войлочными, кожистыми, деревянистыми, мягкомясистыми, могут иметь форму пленок, корочек, могут быть копытообразными или состоять из шляпки и ножки.
Спороносный слой плодового тела —гимений — располагается у более примитивных видов на верхней стороне плодовых тел, а у более высокоорганизованных — на нижней. Гимений базидиальных грибов состоит из базидий с базидиоспорами и парафиз. У некоторых видов в гимении находятся цистиды — крупные клетки, возвышающиеся над гимениальным слоем. Поверхность плодового тела, несущую гимении, называют гименофором. У низших представителей он гладкий, а у более высокоорганизованных имеет форму зубцов, трубочек, пластинок.
Типы гименофоров у грибов:
1. Гладкий. Гладкий гименофор наиболее простой, он не имеет развитой поверхности. Такой тип чаще всего встречается у аскомицетов и находится на верхней поверхности чашевидных апотециев или на наружной поверхности «шляпок» сморчковых.
2. Складчатый (Такой тип имеет поверхность в виде радиальных складок или хаотически расположенных морщин. Встречается в основном у булавовидных и распростёртых плодовых тел, но характерен и для некоторых шляпочных.).
3. Шиповатый (Шиповатый гименофор бывает у распростёртых, кустистых и шляпконожечных плодовых тел. Состоит из конических или игольчатой формы шипиков, расположен обычно на нижней поверхности гриба).
4. Трубчатый. Трубчатый, или пористый слой встречается у болетовых
и трутовиков. Он всегда находится на нижней поверхности шляпки или сидячего плодового тела, представляет собой массу трубочек, открывающихся вниз.
5. Лабиринтовидный. Такой тип спороносного слоя представляет собой модификацию трубчатого с вытянутыми в радиальном направлении порами и отсутствующими в отдельных местах стенками трубочек. Встречается у трутовиковых грибов).
6. Пластинчатый. Пластинки — это выросты ткани гриба на нижней поверхности шляпки, они расходятся радиально от ножки (или от места прикрепления к субстрату сидячей шляпки).
ОТДЕЛ АСКОМИКОТА (ASCOMYCOTA), ИЛИ СУМЧАТЫЕ ГРИБЫ
Лабораторная работа № 5
Цель работы: изучить особенности строения вегетативных и генеративных структур отдела Ascomycota.
1. Познакомиться с особенностями полового процесса сумчатых грибов.
2. Изучить жизненный цикл аскомицетов.
3. Рассмотреть классификацию отдела и познакомиться с отдельными представителями.
Теоретические сведения:
Аскомицеты — грибы с хорошо развитым септированным гаплоидным мицелием. Септы образуются путем инвагинации, т. е. путем впячивания и роста клеточной стенки от периферии к центру. Поэтому в перегородках между клетками имеются поры, через которые перемещаются не только растворенные вещества, но и органеллы. Составными элементами клеточной стенки являются полисахариды хитин и глюканы.
Характерная особенность грибов этого класса – образование после полового процесса сумок, или асков, содержащих 4 – 8 (редко одну) аскоспор (рис. 1). Кроме того, диагностическим признаком сумчатых грибов является наличие ламеллярных двуслойных клеточных стенок с тонким электронноплотным наружным слоем и относительно толстым электронно-прозрачным внутренним слоем. 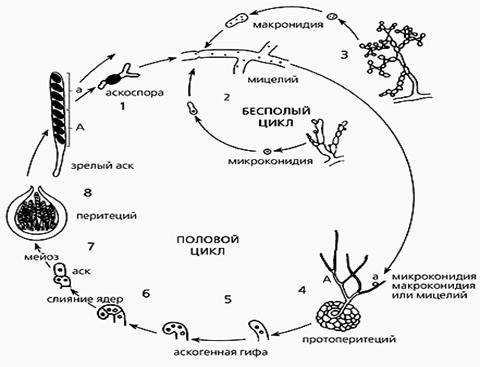
Рис. 1. Половой цикл аскомицетов на примере жизненного цикла Neurospora crassa. Стадии 1-4 — от прорастания аскоспор до оплодотворения — вегетативная фаза; стадии 4-8 — от оплодотворения до созревания.
У базидиальных грибов они многослойные. Вегетативное размножение осуществляется участками мицелия, почкованием или путем расчленения гифы на отдельные клетки и формирования оидий, хламидоспор и т. д.
Важное место в цикле развития аскомицетов играет бесполое размножение при помощи конидий. Отдельные группы грибов могут развивать несколько видов конидиального спороношения.
Типы полового процесса определяются уровнем организации гриба. У примитивных, наиболее просто устроенных аскомицетов (класс Голосумчатые) половой процесс состоит в слиянии содержимого двух клеток (гаметангиев) мицелия и напоминает собой зигогамию или конъюгацию. Образовавшаяся зигота прямо на мицелии превращается в сумку. При этом копуляционное диплоидное ядро без периода покоя зиготы делится редукционно, затем митотически, образуя 8 гаплоидных ядер, которые облекаются цитоплазмой, формируют оболочку и превращаются таким образом в аскоспоры. Сумки у них протуникатные, т. е. с тонкой, слабо дифференцированной оболочкой, которая ослизняется или разрушается при созревании аскоспор. Такая сумка не участвует в рассеивании спор, а является лишь их вместилищем.
У высокоорганизованных аскомицетов (класс Плодосумчатые) формируются женские и мужские органы полового размножения – антеридии и архикарпы (рис. 2, а).
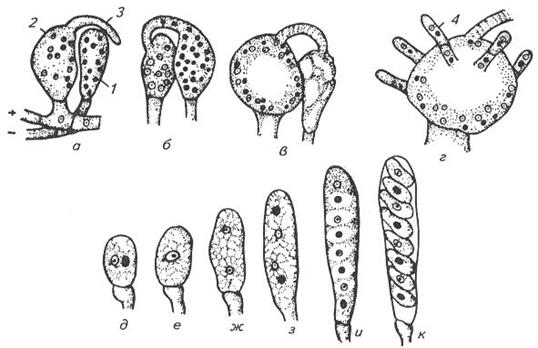
Рис. 2. Половой процесс у сумчатого гриба пиронемы: а – антеридий (1) и архикарп (2) с трихогиной (3); б – переход содержимого антеридия в архикарп, в – образование в аскогоне дикарионов; г – образование дикарионных аскогенных гиф (4); д, е – образование зиготы на верхушке аскогенной гифы; ж, з, и, к – мейоз и развитие сумки с аскоспорами.
Архикарп состоит из нижней расширенной части — аскогона и верхней суженной — трихогины. Содержимое антеридия, недифференцированное на гаметы, перетекает через трихогину в аскогон (рис. 2, б), где происходит плазмогамия, а гаплоидные ядра разного пола сближаются попарно, образуя дикарионы (рис. 2, в). Затем брюшко аскогона прорастает аскогенными гифами, в которых закладываются поперечные перегородки, образующие клетки с одним или несколькими дикарионамн. На концах аскогенных гиф в результате кариогамии с последующим мейозом и митозом образуются сумки с 8 аскоспорами (рис. 2, г, ж, з, и, к). Сумки возникают внутри или на поверхности плодовых тел, образуемых сплетением гиф мицелия.

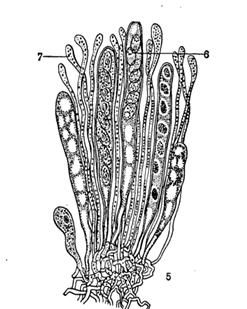
Рис. 3. Типы плодовых тел аскомицетов: 1, 2 – клейстотеции; 3 – перитеции; 4 – апотеции; 5- основание гимениального слоя; 6 – сумка; 7 – парафизы; 8 – перифизы.
Различают три типа плодовых тел: 1) клейстотеции — замкнутые плодовые тела с беспорядочно расположенными внутри них сумками; 2) перитеции имеют отверстие на вершине, а сумки образуются пучком внутри плодовых тел (на дне) и способны по мере созревания активно выбрасывать споры; 3) апотеции — открытые плодовые тела, на поверхности которых палисадным слоем развиваются сумки и бесплодные гифы — парафизы (рис. 3). Совокупность сумок и парафиз составляет гимениальный слой, или гимений. Распространение спор активнее, чем в случае перитеция.
У некоторых грибов формируются ложные плодовые тела — аскостромы, по внешнему виду часто похожие на настоящие перитеции, но отличающиеся от них отсутствием собственного перидия; их оболочкой служит плектенхима стромы.
В цикле развития высших сумчатых грибов закономерно сменяют друг друга три ядерные фазы: длительная гаплоидная, представленная долгоживущим мицелием, на котором развивается конидиальное спороношение, кратковременная дикариотическая – от плазмогамии до кариогамии (существует в виде аскогенных гиф) и самая кратковременная диплоидная (диплоидное ядро материнской клетки сумки) – от кариогамии до мейоза.
Сумчатые грибы — преимущественно сапротрофы на отмерших растительных остатках, почве, пищевых продуктах, различных промышленных материалах и изделиях, вызывают их плесневение и порчу. Некоторые сапротрофы являются продуцентами антибиотиков и других биологически активных веществ. Среди сумчатых грибов встречаются съедобные виды (сморчки, строчки, трюфели) и паразиты, возбудители болезней растений, животных и человека.
Сумчатых грибов насчитывают около 30 000 видов. Аскомицеты подразделяются на 4 класса: Археаскомицеты, Гемиаскомицеты, или Голосумчатые, Настоящие сумчатые и Локулоаскомицеты. Систематика отдела основана на расположении сумок: открыто на мицелии, в особых полостях (локулах) или плодовых телах, а также на особенностях их строения (схема 1).
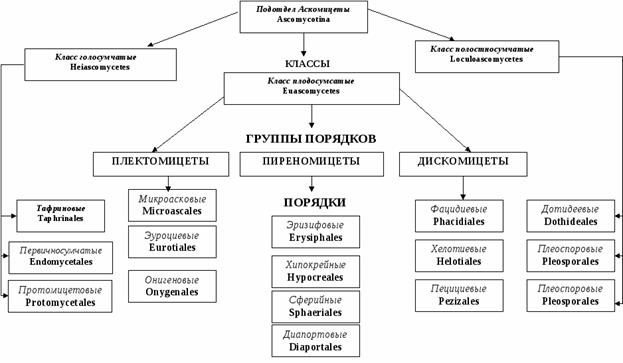
Схема 1. Классификация отдела Ascomycota (на схеме класс Archaeascomycetes внесен в класс Hemiascomycetes).
Класс Археаскомицеты (Archaeascomycetes)
Класс включает одноклеточные и многоклеточные грибы, у которых плодовые тела отсутствуют. Сумки у всех представителей класса эутуникатные, то есть с плотной оболочкой, часто с особым аппаратом для вскрывания и активного распространения спор.
Класс подразделяют на несколько (от 2 до 5) порядков, основным из которых является порядок Тафринальные. Порядок Тафринальные (Taphrinales). К этому порядку относятся исключительно паразиты на высших растениях, у которых они вызывают различные деформации пораженных органов (курчавость листьев, «ведьмины метлы», кармашки плодов, галлообразные вздутия и др.).
Настоящий нитчатый мицелий развивается под кутикулой или в межклетниках, однолетний или зимует в почках и побегах, легко распадается на отдельные членики. Сумки образуются пучками или в виде сплошного гимениального слоя, выступающего на поверхности пораженного органа. Сумки отходят непосредственно от мицелия или каждая помещается на отдельной подсумочной клетке в виде подставки, отделяющейся от сумки поперечной перегородкой. Сумкоспоры шаро-, эллипсовидные или продолговатые, одноклеточные, бесцветные, по 4–8 в сумке, часто в ней почкующиеся. Конидиальное спороношение у них отсутствует.
В их цикле развития наблюдается длительная дикариотическая фаза, а их сумки имеют эутуникатное строение и аскоспоры из них выбрасываются активно. Предполагают, что тафринальные филогенетически связаны с дискомицетами, а отсутствие у них плодовых тел — результат морфологического упрощения в процессе приспособления к паразитизму.
Тафринальные грибы объединены в одно семейство тафриновых (Taphrinophyceae) с одним родом тафрина, включающем около 100 видов, широко распространенных во многих странах мира Северного и Южного полушарий.
Род тафрина (Taphrina) включает грибы с эндофитным мицелием. Сумки образуются непосредственно на мицелии плотным слоем, имеющим сходство с гимением высших сумчатых грибов. Слой сумок обычно имеет желтый, красный, розовый или фиолетовый оттенок. По форме сумки могут быть цилиндрическими, булавовидными или даже удлиненными. Они достигают 60—110 мкм в высоту и 25 мкм в ширину. В основании сумок находится лишенная протопласта клетка, образовавшаяся в результате деления аскогенной клетки и сохраняющая связь сумки с гифами. В сумке 8 аскоспор округлой или яйцевидной формы, 7—10 мкм в диаметре. Они способны к почкованию, в результате чего число их в сумке может увеличиваться в 2—4 раза. Многие из тафриновых грибов гетероталличны.
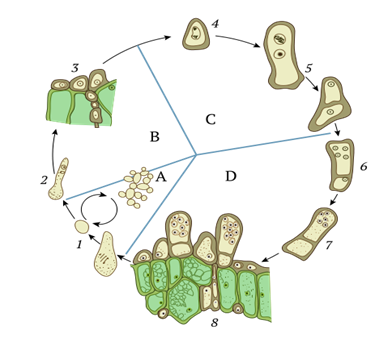
Рис. 4. Жизненный цикл тафрины. А – гаплоидная стадия (дрожжи); В – дикариотическая стадия (мицелий); С – диплоидная стадия (проаски); D – развитие асков и образование половых спор. 1 – аскоспоры и почкующиеся бластоспоры (конидии); 2 – дикариотизация; 3 – дикариотический мицелий на растительной клетке, формирование аскогенного поколения; 4 – кариогамия; 5 – митоз диплоидного ядра, проаски и формирование базальных клеток; 6 – развитие асков после мейоза; 7 – митозы гаплоидных ядер, формирование аскоспор; 8 – формирование спорогенного поколения в растительных клетках.
Наиболее известны 4 вида: тафрина сливовая (Т. pruni) поражает сливу, алычу, вишню, черемуху, вызывая образование так называемых «дутых плодов» или кармашков; тафрина терносливовая (Т. insititiae) вызывает образование «ведъминых метел», волнистость и курчавость листьев слив и тернослив; тафрина тополевая (Т. rhizophora) и тафрина Визнера (Т. wiesneri), паразитирующие соответственно на тополях и вишнях или черешнях.
Класс Гемиаскомицеты, или Голосумчатые (Hemiascomycetes)
Характерный признак класса — отсутствие у грибов аскогенных гиф и плодовых тел, за что они и получили название голосумчатых. Сумки у них протуникатные, развиваются на мицелии из зиготы или аскогенных клеток и расположены беспорядочно, а у одноклеточных — в питательном субстрате. Конидиальное спороношение неизвестно. Половой процесс напоминает зигогамию у мукоральных, однако, в отличие от зигоспоры последних, зигота голосумчатых не переходит в состояние покоя, а сразу же превращается в сумку.
Основной порядок, выделяемый по строению вегетативного тела, образу жизни и особенностям цикла развития – сахаромицетальные.
Порядок Сахаромицетальные (Saccharomycetales). Порядок включает грибы, у которых сумки с 4–8 аскоспорами развиваются одиночно, прямо из зиготы. Встречаются также одно- и многоспоровые сумки. У большинства сахаромицетальных грибов настоящего мицелия нет, а вегетативное тело представлено одиночными клетками, которые размножаются почкованием или реже делением. Некоторые представители имеют хорошо развитый, ветвящийся, септированный мицелий, у которого концевые веточки часто распадаются на дрожжевидные, способные к почкованию клетки. На мицелии сначала развивается бесполое спороношение — многоядерные артроспоры в цепочках (реже удлиненные спорангии), а затем половое — гаметангии, В результате полового процесса образуется зигота, которая превращается в одно- или многоспоровую сумку или прорастает в мицелий.
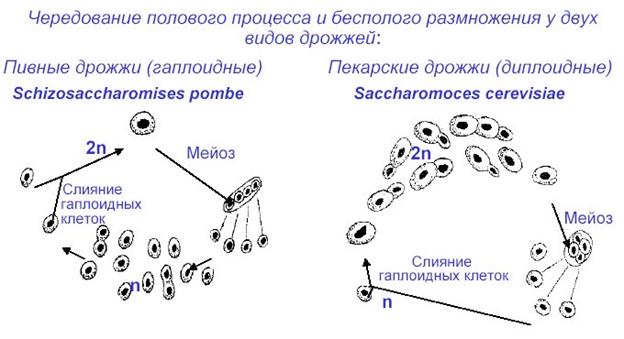
Питаются сахаромицетальные преимущественно сапротрофно, но некоторые паразитируют на животных и человеке.
Род сахаромицес (Saccharomyces) включает грибы, тело которых состоит из отдельных округлых, овальных или удлиненных почкующихся клеток. Иногда образуется псевдомицелий. При почковании могут формироваться разветвленные короткие цепочки. При неблагоприятных условиях возникают эллипсовидные, шаро- или булавовидные артроспоры или 1–4-споровые сумки с округлыми, гладкими, бесцветными аскоспорами.
Описано более 50 видов и несколько десятков разновидностей, широко распространенных по всему земному шару. К роду относятся так называемые культурные дрожжи, т. е. виды и расы, получившие широкое применение в различных отраслях пищевой промышленности благодаря их способности к активному сбраживанию сахаров с образованием наибольшего по сравнению с другими дрожжами количества спирта (от 10 до 19% по объему). Из них наибольшее практическое значение имеют хлебные, или пекарские (S. cerevisiae), пивные (S. carsbergensis и S. uvarum) и винные (S. vini) дрожжи.
Пекарские дрожжи в природных условиях не встречается, а винные можно встретить в природе там, где имеются сбраживаемые ими углеводы: на поверхности ягод винограда, фруктов, в нектаре цветков, слизетечениях берез, кленов и других деревьев.
Задания
1. Для приготовления препарата тафрины необходимо иметь фиксированные в спирте пораженные листья или плоды персика, вишни, слив или черемухи. Сделать поперечные срезы через пораженные ткани. Срезы рекомендуется помещать в 5—10%-ный раствор КОН. На поверхности среза виден слой сумок на разных стадиях созревания. В зрелых сумках содержится по 8 овальных бесцветных спор. Иногда можно наблюдать их почкование. Зарисовать сумки со спорами и хламидоспоры. (Под сумкой хорошо заметна клетка — ножка, пустая клетка мицелия или хламидоспора, вырост которой развивается в сумку.)
2. Кончиком стеклянной палочки небольшое количество дрожжевой массы перенести в каплю воды на предметном стекле и, закрыв препарат покровным стеклом, рассмотреть его при большом увеличении микроскопа. Клетки хлебных дрожжей имеют округло-эллипсовидную форму (одиночные или соединенные по 2–4 в цепочки), а винные – в виде ветвистых колоний, состоящих из цилиндрических клеток. На многих клетках можно видеть маленькие выпуклины — почки. В мелкозернистом содержимом живых клеток дрожжей хорошо заметны крупные, прозрачные вакуоли, занимающие обычно центральное положение. Зарисовать клетки дрожжей обоих видов.
Источники:
http://gendocs.ru/v6423/%D0%BB%D0%B5%D0%BA%D1%86%D0%B8%D0%B8_%D0%BF%D0%BE_%D1%84%D0%B8%D1%82%D0%BE%D0%BF%D0%B0%D1%82%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D0%B8?page=6
http://worldofschool.ru/biologiya/stati/miko/griby/askomik/otdel-sumchatye-griby-ili-askomikota-ascomycota
http://studopedia.su/1_6474_otdel-sumchatie-gribi-ili-askomitseti-ASCOMYCOTA.html
http://lektsii.org/12-36756.html

